■今更メタロチオネインなのかと言われそう。
そう、しつこいよ(笑)

メタロチオネインって、
鉄にも反応するけど結びつかない、なんて性質がある。
鉄が出る時ってどんな時?何の為
鉄はトランスフェリンで運ばれフェリチンに貯蓄する鉄代謝がある。
そしてフェリチンは色んなものを拾い集める。セシウムもね。ラジウムもね。カドミウムもね。

■
実はフェリチン→、メタロチオネイン→代謝の流れがあるんじゃないだろうか??
もちろんメタロチオネイン直接の時もある。
そうしてそのメタロチオネインは
どこで分解されるのかな??
下によれば、
膵臓や肝臓に多く蓄積する。
セシウムに限らなくても沢山の核種がある。
もちろん、必要なものと間違えて取り込むケース(ストロンチウム)も怖い
でも、重金属として、もし生化学的に処理されたら・・・
これは一つの可能性。
でも、原発ができてから膵臓がんが増加した、なんて話もあった。
糖尿病も増加の一歩を辿っている、とか?
そう、原爆症とか、放射線障害じゃなくて、臓器障害=成人病
として実は放射性物質は影響を及ぼす可能性が否定できない
統計的には怪しいちゃあ怪しいんだけどね。
そう、このエントリーを見ると
近未来が書いているのかもしれない
2011年06月18日
【これぞ統計マジック】放射能と健康【危ないのは本当】
気をつけるべきは肝臓がんとすい臓がん、
下手をすると糖尿病も?・・・
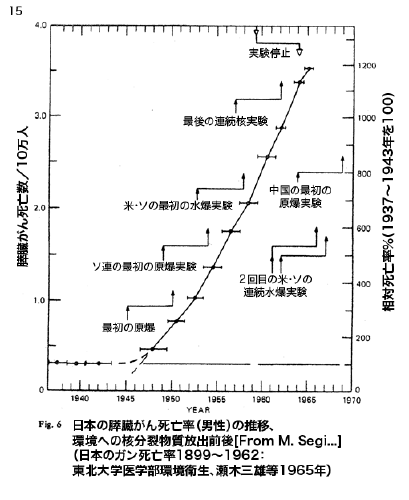
特に子供に、
エビオスをお勧めしきれないのはこれが理由・・・
放射性重金属取込みが多い場合、
メタロチオネインの代謝経路で
沈黙の臓器2個がターゲットとなるかもしれないから
(外部放射線=ラジカル除去、に対しては非常に強力だと思います)
ラクトフェリンは問題ないと思います^^
取り込んだ時には、人の体は排出能力に限りがあります。
膵、肝、腎
これらは紛れもなく消耗品です。
もしお子さんがいるならば、出来るだけ労わって頂ければと思います。
そしてその方法は出来るだけ放射性物質を摂取しない事。
ま、40過ぎた大人はじゃんじゃん食ってよし。
もちろんメタロチオネイン以外を使う経路もあるかもしれない。
排出は膵、肝、腎、がその多くに関る以上、
やっぱり労わって欲しいものですね。
但し、そういった被害に合うのはごく一部の運の悪い人、
統計のカラクリを理解した上で悩んでくださいませ。
********************************************
これは
徳島文理大学のメタロチオネインのページ
(1)メタロチオネインの構造
メタロチオネイン(Metallothionein)は分子量が約6500と低分子量の金属結合たんぱく質である。metallo-は金属、thio-は多く含まれるシステインのthol-(SH)基、teinは蛋白(protein)から由来している。
構成アミノ酸は生物種により異なるが約61個のアミノ酸残基から構成され、その1/3をシステインが占めるという珍しいタンパク質であり、チロシン、フェニルアラニン、トリプトファンなどの芳香族アミノ酸を含まない。そのためカドミウム、亜鉛をあわせて7個の金属がSH基と結合をしている。銅や銀は12個結合する。
生体内存在形:
2つのクラスターを形成(αクラスター、βクラスター)する事により、1分子当たり亜鉛、Cd7原子と結合する。2つのクラスターへの金属の取り込みには規則性があり、αクラスターに4原子が取り込まれた後にβクラスターに3原子の金属が捕捉される。
酸化還元状態でみると、分子内システインのSH基はすべて還元型で酸化型(S-S結合)はなく、システイン残基が重金属解毒、金属代謝調節やラジカル消去などの生理機能作用発現に役割を担っている。
たんぱく質については、亜鉛結合たんぱく質を探索していたハーバード大のVallee教授のもとでKagiおよびMargoshes博士が1975年に馬の腎臓からカドミウムと亜鉛を多量に含有するたんぱく質として単離し、さらに詳細な解析データを1960年に報告した。1976年にアミノ酸配列決定、1980年には遺伝子クローニングが行われている。
(2) 存在
地球上には、
アカパンカビ、かになどの海産生物から哺乳類に至るまで基本的構造を保存して分布している。
人体中濃度は組織や金属曝露状況に依存するが、ヒト血清中では数ng/mlで検出は困難である。
(3) 機能解析
遺伝子発現欠損マウス作製:殆どの組織で恒常的に発現しているが
1993,1994年に作製された遺伝子発現欠損マウスが致死に至ることはなかったが、カドミウムなど環境からの刺激には低濃度曝露により致死に至る(1)。金属によってその毒性は異なっている。
(4) 吸収・代謝・排泄 体内および組織内分布
メタロチオネインはI型からIV 型まで存在する。
I、II型は肝臓、腎臓、膵臓、心臓などほとんどの組織に分布している。III型は特に脳に局在するが、小腸、膵臓、舌、腎臓などでも検出される。IV型は扁平上皮細胞にみられるが詳細は不明である。
I型、II型アイソフォームの金属との親和性はZn < Cd < Cu < Hg, Agの順である。
細胞内ではメタロチオネインはその大半は通常細胞質に存在し、再生肝では核に移行し増殖制御作用が推察されたが、最近になって、メタロチオネインが欠損すると肝再生がおこらない事が示された(2)。また、メタロチオネインがミトコンドリアの内膜と外膜の間に局在し、ミトコンドリア酸素呼吸を調節することが報告された(3)。酸素呼吸の場であるミトコンドリアは、細胞内で生成される活性酸素の大半を恒常的に産生している事から強力な活性酸素消去能を持つメタロチオネインの役割が注目される。
臓器特異性:
ヒトでは加齢に伴いカドミウムが腎組織に高濃度に蓄積する事に伴い、日本人は特に高い。膵臓や肝臓に多く蓄積する。代謝の早さを示す生物学半減期は結合金属により異なり、ラット肝臓では亜鉛結合型は約20時間、銅結合型は16時間に比べカドミウム結合型は4〜5日と長い。
(5) 生理作用
金属の組織維持機能:メタロチオネインが欠損するとは亜鉛やカドミウムが組織に取り込まれた後に濃度は減少するので、メタロチオネインは金属を組織に維持する機能を有している。
重金属毒性の解毒作用:カドミウムや水銀などを予め投与しメタロチオネイン誘導後に、多量の金属を投与しても毒性発現が抑えられ、これらの重金属の毒性を解毒する作用を持っている。メタロチオネインが欠損するとカドミウム毒性発現は大きい(1)。
亜鉛や銅などの必須金属の代謝調節機能:一時的に多量組織に入ってきた亜鉛や銅などの必須金属を貯蔵し、必要に応じて分布する代謝調節する。
ラジカル消去作用:SH基を多く含むので活性酸素を含むラジカル消去作用を示す(4)。生体内ではスーパーオキシドが生成すると過酸化水素に変換し、さらに水に分解する防御酵素が存在する。活性酸素の毒性は過酸化水素が鉄や銅イオンの存在下にヒドロキシラジカルを生成するが、それに対してメタロチオネインは生体物質の中で最も強いグルタチオンと比べても約400倍の親和性を有している。
生体内では呼吸に伴い、細胞のミトコンドリアで常に活性酸素が生成することに対してメタロチオネインは障害を抑制しているが、細胞内小胞体でおこるストレスに対しても作用機構は不詳であるが抑制する作用を示す事を我々は見いだしている。
(6) 欠乏症
亜鉛欠乏症では合成が不十分で組織濃度は低下する。
(7) 過剰症
ウイルソン病で組織に銅が蓄積しメタロチオネインに結合しているが、病気が進展し大量に蓄積した銅-メタロチオネインから銅を介したラジカルが産生されて組織障害を引き起こす。
(8) タンパク質・酵素・天然物
メタロチオネインは酵素作用を有しないが、赤パンかびから、かになどの海洋生物、マウスや人などの哺乳類まで広範な生物組織に存在する。アミノ酸配列は基本的に保存されている。
メタロチオネインは種々の要因により誘導され新たに合成されるので様々な生物現象と関わる多彩な機能が推定されている。重金属により誘導される事を基本として、ストレスや炎症に早期に応答する蛋白質である。
亜鉛、銅の必須金属やカドミウム、無機水銀などの有害金属およびグルココルチコイドによりメタロチオネインたん白が新たに作られる。それ以外のストレスや炎症ではインターロイキンー6などのサイトカインを介して間接的に合成される場合が多い。
表1 メタロチオネインを誘導する代表的要因
1.必須金属 亜鉛、銅
2.有害金属 カドミウム、無機水銀
3.生理活性物質 サイトカイン(インターロイキンー6、腫瘍壊死因子)(5)
4.ホルモン グルココルチコイド、レプチン(6)
5. 起炎剤 エンドトキシン、イソプロパノール、エタノール、四塩化炭素
6.ストレス 拘束ストレス、絶食、強制運動、小胞体ストレス誘導剤(ツニカマイシン)(7)
7.物理的ストレス 紫外線照射、高圧酸素曝露、x線照射
8.活性酸素産生剤 アンチマイシンA、2、4--ジニトロフェノール、ストレプトゾトシン、パラコート
(9) 定量法
メタロチオネインは金属結合たんぱく質なので、大別して(i)たんぱく質部分(thionein)に注目した抗体による検出や(ii)金属結合性を利用し、カドミウムや水銀でメタロチオネイン結合金属を置換し、その金属量から算出する方法がある。
(10) 総説
鈴木和夫. メタロチオネインが関与する微量金属の毒性と疾患. 化学総説 1995 ; 27 : 153-165.
Davis SR, Cousins RJ. Metallothionein expression in animals: A physiological perspective on function. J Nutr
2000 ; 130 : 1085-1088.
Andrews GK. Regulation of metallothionein gene expression by oxidative stress and metal ions. Biochem
Pharmacol 2000 ; 59 : 95-104.
Sato M., Kondoh M., Recent studies on metallothionein:Protection against toxicity of heavy metals and oxygen free radicals, Tohoku J. Exp.Med., 196,9-22,2002
近藤昌夫、佐藤政男、生体防御にかかわる蛋白質:メタロチオネイン、日本衛生学会誌、56、634−640,2002
Haq F. Mahoney M., Koropatnick J., Signaling events for metallothionein indeuction, Mutation Res., 533,211-226,2003
(11) 文献
Park JD, Liu Y, Klaassen CD., Protective effect of metallothionein against the toxicity of cadmium and other metals(1), Toxicology. 2001 Jun 21;163(2-3):93-100.
Oliver JR, Mara TW, Cherian MG, Impaired hepatic regeneration in metallothionein-I/II knockout mice after partial hepatectomy, Exp Biol Med (Maywood).;230(1):61-7, 2005
Ye B, Maret W, Vallee BL. Zinc metallothionein imported into liver mitochondria modulates respiration. Proc Natl Acad Sci USA 2001 ; 98 : 2317-2322.
Zhou Z, Wang L, Song Z, Saari JT, McClain CJ, Kang YJ., inc supplementation prevents alcoholic liver injury in mice through attenuation of oxidative stress, Am J Pathol., 166(6):1681-90, 2005
Sato M, Sasaki M, Hojo H. Antioxidative roles of metallothionein and manganese superoxide dismutase induced by tumor necrosis factor-α and interleukin-6. Arch Biochem Biophys 1995 ; 316 : 738-744.
Waelput W, Verhee A, Broekaert D, Eyckerman S, Vandekerckhove J, BeattieJH, Tavernier J、
Identification and expression analysis of leptin-regulated immediate earlyresponse and late
target genes、Biochem J. 348 Pt 1:55-61, 2000.
Kondoh M, Tsukada M, Kuronaga M, Higashimoto M, Takiguchi M, Himeno S, Watanabe Y, Sato M.、Induction of hepatic metallothionein synthesis by endoplasmic reticulum stress in mice、Toxico。Lett.; 148(1-2):133-9, 2004
 【たばこ】喫煙しませんが、一応大丈夫、もう一つくらいバイアスがいります
【たばこ】喫煙しませんが、一応大丈夫、もう一つくらいバイアスがいります 
















